¿Qué es la fase oscura de la fotosíntesis?
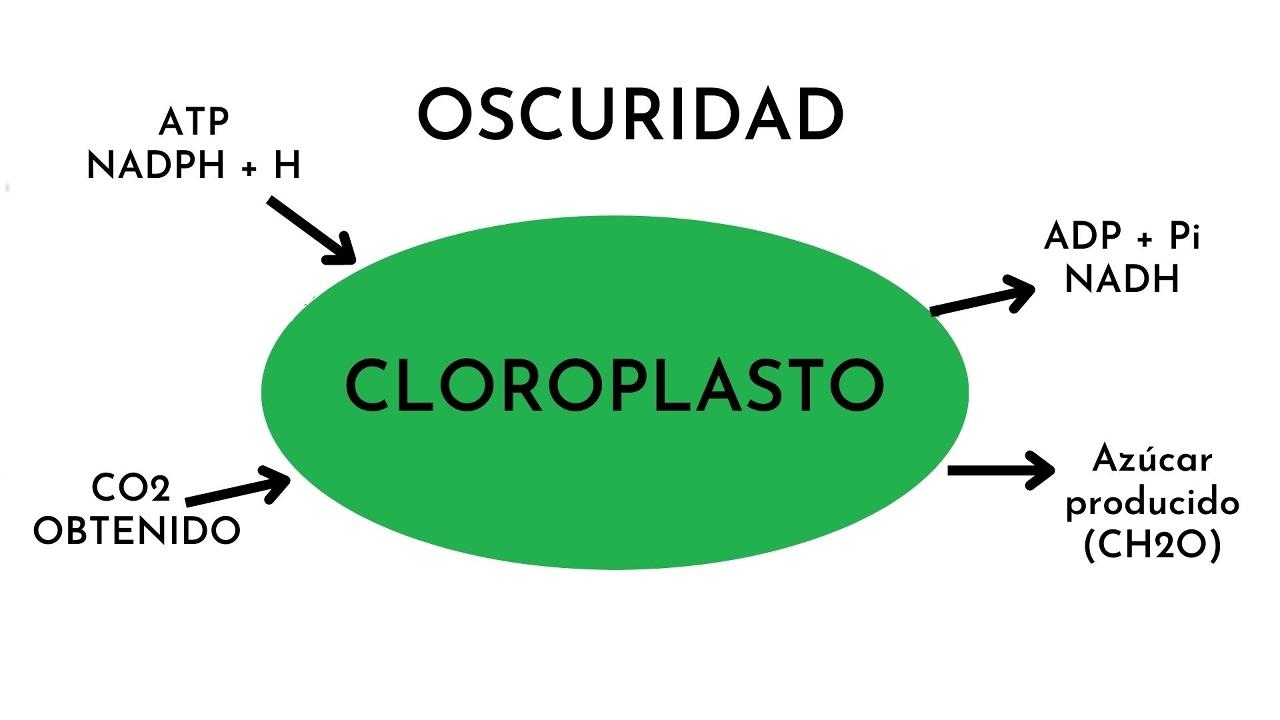
La fase oscura de la fotosíntesis es el proceso bioquímico mediante el cual las plantas convierten el dióxido de carbono en glucosa. Se le dice oscura porque no se necesita la luz para el proceso. También se conoce como fase de fijación de carbono o ciclo de Calvin-Benson. Este proceso ocurre en el estroma del cloroplasto.
En la fase oscura, la energía química la suministran los productos generados en la fase luminosa. Estos productos son las moléculas energéticas ATP (trifosfato de adenosina) y el NADPH (un portador de electrones reducido).
La materia prima fundamental para el proceso en la fase oscura es el carbono, que se obtiene a partir del dióxido de carbono. El producto final son los carbohidratos o azúcares simples.
Estos compuestos de carbono que se obtienen, son la base fundamental de las estructuras orgánicas de los seres vivos.
Características de la fase oscura de la fotosíntesis
– Se denomina oscura por el hecho de no requerir para su desarrollo la participación directa de la luz solar. Este ciclo ocurre durante el día o la noche.
– La fase oscura se desarrolla principalmente en el estroma del cloroplasto en la mayoría de los organismos fotosintéticos. El estroma es la matriz que llena la cavidad interna del cloroplasto en torno al sistema de tilacoides (donde se lleva a cabo la fase luminosa).
– En el estroma se encuentran las enzimas necesarias para que ocurra la fase oscura. La más importante de estas enzimas es la rubisco (ribulosa bifosfato carboxilasa/oxigenasa), la proteína más abundante, representando entre el 20 al 40% de todas las proteínas solubles existentes.
Mecanismos
El carbono requerido para el proceso se encuentra en forma de CO₂ (dióxido de carbono) en el ambiente. En el caso de algas y cianobacterias, el CO₂ está disuelto en el agua que las rodea. En el caso de las plantas, el CO₂ llega a las células fotosintéticas a través de los estomas (células epidérmicas).
Ciclo de Calvin-Benson
Este ciclo tiene varias reacciones:
Reacción inicial
El CO₂ se fija en un compuesto aceptor de cinco carbonos (ribulosa 1,5-bifosfato o RuBP). Este proceso es catalizado por la enzima rubisco. El compuesto resultante es una molécula de seis carbonos.
La misma se rompe rápidamente y forma dos compuestos de tres carbonos cada uno (3-fosfoglicerato o 3PG).
Segundo proceso
En estas reacciones se utiliza la energía proporcionada por el ATP proveniente de la fase lumínica. Ocurre una fosforilación impulsada por la energía del ATP y un proceso de una reducción mediado por el NADPH. Así, el 3-fosfoglicerato es reducido a gliceraldehído 3-fosfato (G3P).
El G3P es un azúcar de tres carbonos fosfatada, también llamada triosa fosfato. Solo una sexta parte del gliceraldehído 3-fosfato (G3P) se transforma en azúcares como producto del ciclo.
Este metabolismo fotosintético se denomina C3, porque el producto básico que se obtiene es un azúcar de tres carbonos.
Proceso final
Las partes del G3P que no se transforman en azúcares, es procesada para formar ribulosa monofosfato (RuMP). La RuMP es un producto intermedio que se transforma en ribulosa 1,5-bifosfato (RuBP). De esta forma, se recupera el aceptor de CO₂ y se cierra el ciclo de Kelvin-Benson.
Del total de RuBP producido en el ciclo en una hoja típica, solo un tercio se convierte en almidón. Este polisacárido se almacena en el cloroplasto como fuente de glucosa.
Otra parte es convertida en sacarosa (un disacárido) y transportada a otros órganos de la planta. Posteriormente, la sacarosa es hidrolizada para formar monosacáridos (glucosa y fructuosa).
Otros metabolismos fotosintéticos
En condiciones ambientales particulares, el proceso fotosintético de las plantas ha evolucionado y se ha hecho más eficiente. Esto ha llevado a la aparición de rutas metabólicas distintas para la obtención de azúcares.
Metabolismo C4
En ambientes cálidos, los estomas de la hoja se cierran durante el día para evitar la pérdida de vapor de agua. Por lo tanto, la concentración de CO₂ en la hoja disminuye en relación con la de oxígeno (O2). La enzima rubisco posee una doble afinidad de sustrato: CO₂ y O2.
A bajas concentraciones de CO₂ y altas de O2, la rubisco cataliza la condensación de O2. Este proceso se denomina fotorrespiración, y disminuye la eficiencia fotosintética. Para contrarrestar la fotorrespiración, algunas plantas de ambientes tropicales han desarrollado una anatomía y fisiología fotosintética particular.
Durante el metabolismo C4, el carbono se fija en las células del mesofilo y el ciclo de Calvin-Benson ocurre en las células de la vaina clorofiliana. La fijación del CO₂ ocurre durante la noche. No ocurre en el estroma del cloroplasto, sino en el citosol de las células del mesofilo.
La fijación del CO₂ ocurre por una reacción de carboxilación. La enzima que cataliza la reacción es el fosfoenolpiruvato carboxilasa (PEP-carboxilasa), que no es sensible a bajas concentraciones de CO₂ en la célula.
La molécula aceptora de CO₂ es el ácido fosfoenolpirúvico (PEPA). El producto intermedio obtenido es el ácido oxaloacético u oxalacetato. El oxalacetato es reducido a malato en algunas especies de plantas o a aspartato (un aminoácido) en otras.
Posteriormente, el malato se traslada a las células de la vaina fotosintética vascular. Aquí es descarboxilado y se produce piruvato y CO₂.
El CO₂ entra al ciclo de Calvin-Benson y reacciona con la rubisco para formar PGA. Por su parte, el piruvato retorna a las células de mesofilo, donde reacciona con ATP para regenerar el aceptor de dióxido de carbono.
Metabolismo CAM
El metabolismo ácido de las crasuláceas (CAM, por sus siglas en inglés) es otra estrategia para la fijación del CO₂. Este mecanismo ha evolucionado de forma independiente en diversos grupos de plantas suculentas.
Las plantas CAM usan tanto la vía C3 como la C4, tal como ocurre en las plantas C4. Pero la separación de ambos metabolismos es temporal.
El CO₂ es fijado en la noche mediante la actividad de la PEP-carboxilasa en el citosol y se forma oxalacetato. El oxalacetato es reducido a malato, que se almacena en la vacuola como ácido málico.
Posteriormente, en presencia de la luz, el ácido málico es recuperado de la vacuola. Es descarboxilado y el CO₂ se transfiere a la RuBP del ciclo de Calvin-Benson dentro de la misma célula.
Las plantas CAM tienen células fotosintéticas con grandes vacuolas donde se almacena el ácido málico, y cloroplastos donde el CO₂ obtenido del ácido málico es transformado en carbohidratos.
Productos finales
Al finalizar la fase oscura de la fotosíntesis, se producen distintos azúcares. La sacarosa es un producto intermedio que es rápidamente movilizado de las hojas a otras partes de la planta. Se puede utilizar directamente para la obtención de glucosa.
El almidón es utilizado como sustancia de reserva. Puede acumularse en la hoja o ser transportado a otros órganos, como tallos y raíces. Ahí se mantiene hasta que es requerido en distintas partes de la planta. Es almacenado en unos plastidios especiales, denominados amiloplastos.
Los productos obtenidos de este ciclo bioquímico son vitales para la planta. La glucosa producida es empleada como fuente de carbono para constituir compuestos, tales como aminoácidos, lípidos o los ácidos nucleicos.
Por otra parte, los azúcares producto de la fase oscura generados representan la base de la cadena alimenticia. Estos compuestos representan paquetes de energía solar transformada en energía química, utilizados por todos los organismos vivos.
Referencias
- Raven, P.H., R.F. Evert y S.E. Eichhorn (1999). Biology of Plants. WH Freeman and Company Worth Publishers.
- Solomon, E.P., L.R. Berg y D.W. Martin (2001). Biología. McGraw-Hill Interamericana.