
¿Qué es el dominio Archaea?
El dominio Archaea o reino Archaea representa uno de los tres dominios de la vida. Está compuesto por organismos procariotas unicelulares microscópicos muy similares y al mismo tiempo muy diferentes a las bacterias y a los eucariotas en muchos aspectos.
La existencia de este grupo fue demostrada hace poco tiempo, más o menos a finales de 1970, por un grupo de investigadores liderados por Carl Woese, quienes consideraron que la vida podía dividirse en eucariotas y en dos tipos de organismos procariotas: las bacterias y las arqueas, también llamadas arqueobacterias.
Los estudios de Woese y colaboradores estuvieron basados en análisis filogenéticos comparativos entre las secuencias de los ARN ribosomales de todos los seres vivos, y la noción de los tres dominios se mantiene, a pesar de la inmensa cantidad de secuencias que se continúa añadiendo a los catálogos de ARN ribosomal (bases de datos).
Dichos estudios consiguieron vislumbrar la posibilidad de que las arqueas sean un grupo hermano de los eucariotas, a pesar de sus similitudes con sus homólogos procariotas (las bacterias), por lo que podrían representar el “eslabón perdido” entre procariotas y eucariotas.
Se cree que estos microorganismos representan más del 20% de todos los procariotas existentes en las aguas oceánicas, entre el 1 y el 5% en los suelos, y el grupo más dominante en los sedimentos marinos y los hábitats geotérmicos.
Las arqueas se caracterizan por habitar en condiciones “extremas” como aguas termales, salinas, ambientes con temperaturas muy elevadas y pH muy ácido, sitios inhóspitos donde la concentración de oxígeno es muy reducida o nula, etc.
Origen evolutivo y relaciones filogenéticas
Existen tres teorías acerca del posible origen de los tres dominios de la vida propuestos por Woese y colaboradores:
- Las bacterias divergieron primero, formando un linaje que produjo a las arqueas y a los eucariotas
- Un linaje “proto-eucariótico” divergió a partir de un linaje completamente procariótico (el de las bacterias y las arqueas)
- Las arqueas divergieron de un linaje que posteriormente dio lugar a los eucariotas y a las bacterias
En 1989, dos investigadores, Gogarten e Iwabe, propusieron independientemente una forma de analizar filogenéticamente a todos los organismos vivos (cosa imposible de hacer a partir de estudios de secuencias de genes únicos).
Iwabe se valió del análisis de las secuencias de genes producidos en eventos “tempranos” de duplicación génica, “enraizando” el árbol de la vida a partir de la comparación de la secuencia de los genes parálogos que codifican factores de elongación.
Los factores de elongación son proteínas de unión a GTP que participan en la traducción, específicamente en la unión de moléculas de ARN de transferencia amioacilados a los ribosomas y en la translocación del peptidil ARN de transferencia.
De acuerdo con los resultados de las comparaciones entre las secuencias de los tres grupos, los genes que codifican para los factores de elongación en las arqueas son más parecidos a los de los organismos eucariotas que a los de las bacterias.
Gogarten, por otra parte, comparó las secuencias de otros genes producidos por eventos de duplicación, concretamente de aquellos que codifican para las subunidades tipo V y tipo F de la enzima ATPasa que se encuentran en arqueas/eucariotas y en bacterias, respectivamente.
Los resultados obtenidos por Gogarten, al igual que los señalados anteriormente, demuestran que estos genes en las arqueas (que presuntamente surgieron por eventos de duplicación genética) están más estrechamente emparentados con los eucariotas que con sus homólogos bacterianos.
Estos argumentos fueron apoyados también por el análisis conducido por otros investigadores muchos años después, quienes emplearon secuencias de otra familia de genes duplicados, la de las ARN de transferencia aminoacil sintetasas, fortaleciendo la concepción de la “cercanía” entre arqueas y eucariotas.
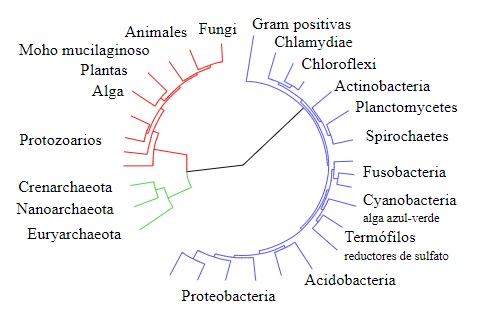
El árbol de Woese
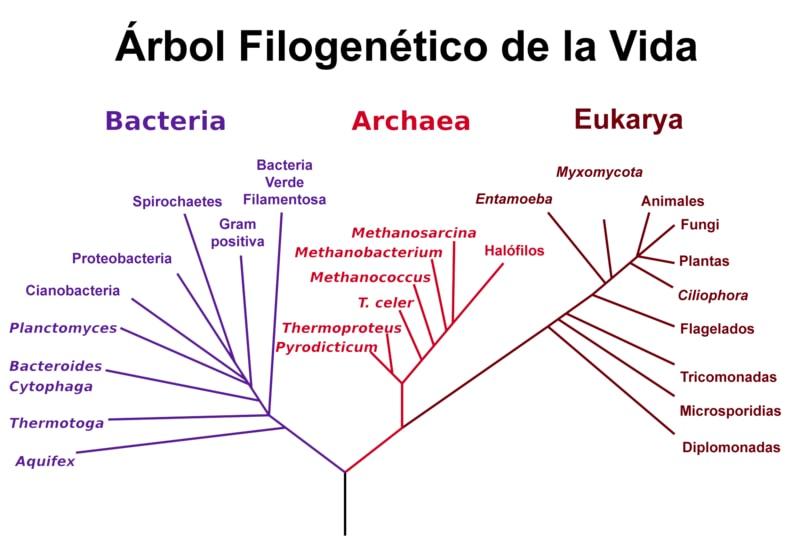
Woese empleó los análisis realizados por Gogarten e Iwabe y otros estudios relacionados con las secuencias de ARN ribosomal para proponer su “versión” del árbol de la vida, donde se hace evidente que las arqueas y los eucariotas constituyen grupos “hermanos”, separados de las bacterias a pesar de que las secuencias de ARN ribosomal entre arqueas y bacterias son más parecidos entre sí.
Características del dominio Archaea
Las arqueas son conocidas por algunas características muy particulares (propias) y también por presentar “combinaciones” de características que alguna vez se pensó eran exclusivas para las bacterias o para los organismos eucariotas.
– Al igual que las bacterias, las arqueas son organismos procariotas, es decir, que en su interior el material genético no está encerrado por una membrana (no tienen núcleo) y no existen orgánulos membranosos citoplasmáticos.
– Generalmente son microorganismos de tamaño similar al de las bacterias, su ADN se encuentra en la forma de un cromosoma circular y algunos fragmentos circulares de menor tamaño conocidos como plásmidos.
– Comparten con las bacterias la presencia de enzimas topoisomerasas y girasas similares, lo que representa una evidencia “indirecta” de la “cercanía” en cuanto a la estructura cromosómica de ambos grupos de organismos.
– Sin embargo, los genes de las arqueas muestran una gran homología con muchos genes eucariotas, especialmente con aquellos cuyo descubrimiento deriva de estudios con antibióticos.
– La maquinaria de replicación, transcripción y traducción de eucariotas y arqueas es muy parecida, sobre todo respecto a la enzima ADN polimerasa.
– Sus genes codificantes para proteínas no tienen intrones (pero otros sí), caso contrario a los genes eucariotas. Además, las arqueas tienen proteínas similares a las histonas asociadas con su ADN, presentes en eucariotas y ausentes en bacterias.
– Se caracterizan por la presencia de éter-lípidos de isoprenil en sus membranas celulares, así como por la ausencia de lípidos acil-éster y de un ácido graso sintetasa.
– Una de las subunidades de su enzima ARN polimerasa está dividida y sus ARN mensajeros, así como en las bacterias, no poseen “capuchas” (del inglés cap) en sus extremos 5´.
– Tienen un rango muy específico de sensibilidad a antibióticos y poseen enzimas de restricción tipo II muy parecidas a las que se han descrito para las bacterias.
– Otra característica importante tiene que ver con el hecho de que gran parte de las arqueas poseen pared celular, pero a diferencia de las bacterias, esta no está compuesta por peptidoglicano.
Características de sus lípidos de membrana
Los lípidos de membrana de las arqueas difieren considerablemente de los que se encuentran en las bacterias y en los organismos eucariotas, y esto ha sido considerado como una característica diferencial muy importante.
La principal diferencia entre estas moléculas anfipáticas (con un extremo polar hidrofílico y uno apolar hidrofóbico) es que la unión entre la porción de glicerol y las cadenas de ácidos grasos en los lípidos de arqueas es a través de un enlace éter, mientras que en las bacterias y los eucariotas corresponde a un enlace éster.
Otra diferencia importante es que las arqueas poseen lípidos con ácidos grasos caracterizados por la presencia de cadenas isoprenil altamente ramificadas con grupos metilo, entretanto los eucariotas y las bacterias poseen predominantemente ácidos grasos de cadenas no ramificadas.
Los lípidos de los eucariotas y de las bacterias están “construidos” sobre un esqueleto de glicerol al cual están esterificadas las cadenas de ácidos grasos en las posiciones correspondientes a los átomos de carbono 1 y 2, pero en las arqueas los éteres de glicerol contienen ácidos grasos en las posiciones 2 y 3.
Otra diferencia respecto a los lípidos de membrana tiene que ver con su ruta biosintética, pues algunas enzimas también son diferentes en las arqueas.
Por ejemplo, algunas especies de arqueas poseen una enzima prenil transferasa bifuncional, que se encarga de aportar los precursores tanto para la síntesis del escualeno como para la síntesis de los isoprenoides de gliceril-lípidos. En las bacterias y los eucariotas estas funciones son llevadas a cabo por enzimas separadas.
Clasificación de las arqueas
De acuerdo con los datos de las secuencias de las subunidades pequeñas de los ARN ribosomales de las arqueas, este grupo ha sido dividido principalmente en dos “filos”, los cuales se conocen como el filo Crenarchaeota y el filo Euryarchaeota, cuyos miembros son, sobre todo, arqueas cultivadas in vitro.
No obstante, muchas de las arqueas descritas recientemente no han sido cultivadas in vitro y solo están lejanamente relacionadas con las secuencias que han sido aisladas de las especies mantenidas en los laboratorios.
Filo Crenarchaeota
Este grupo está formado principalmente por especies de arqueas hipertermófilas y termoacidófilas, es decir, aquellos géneros de arqueas que habitan ambientes inhóspitos con condiciones térmicas y de pH extremas.
Está compuesto por una sola clase taxonómica, que se conoce como Thermoprotei, dentro de la cual se encuentran los siguientes cinco órdenes taxonómicos: Acidilobales, Desulfurococcales, Fervidicoccales, Sulfolobales y Thermoproteales.
Ejemplo de algunos de los géneros pertenecientes a dichas clases pueden ser los géneros Sulfolobus, Desulforococcus, Pyrodictium, Thermoproteus y Thermofilum.
Filo Euryarchaeota
Los miembros de este grupo tienen un rango ecológico un poco más amplio, pudiendo hallarse en él algunas especies hipertermófilas, metanógenas, halófilas e incluso metanógenas termofílicas, arqueas desnitrificantes, reductoras de azufre, oxidadoras de hierro y algunas organotrofas.
Las clases taxonómicas descritas para los euriarqueotas son ocho y se conocen como Methanopyri, Methanococci, Methanobacteria, Methanomicrobia, Archaeglobi, Halobacteria, Thermococci y Thermoplasmata.
Muchas de las arqueas pertenecientes a este grupo están ampliamente distribuidas, hallándose en los suelos, sedimentos y aguas marinas, así como en los ambientes extremos descritos.
Filo Thaumarchaeota

Este filo fue definido hace relativamente poco tiempo y muy pocas de las especies pertenecientes al mismo han sido cultivadas in vitro, por lo que se sabe poco de estos organismos.
Todos los miembros del filo obtienen su energía a partir de la oxidación de amonio y están distribuidos globalmente en cuerpos de agua dulce, suelos, sedimentos y aguas termales.
Filos Korarchaeota, Aigarchaeota y Goarchaeota

Algunos investigadores expertos en la materia, basándose en el análisis de secuencias genómicas, han determinado recientemente la existencia de tres filos adicionales en el reino Archaea, aunque las especies propuestas para estos filos todavía no fueron aisladas en un laboratorio.
En general, los miembros pertenecientes a estos filos han sido hallados bajo la superficie de muchos ecosistemas terrestres y marinos, pero también en aguas termales y en sistemas hidrotermales del mar profundo.
Nutrición
La mayoría de las especies de arqueas con organismos quimiótrofos, es decir, que son capaces de emplear compuestos inorgánicos altamente reducidos para conseguir la energía que necesitan para “mover” su maquinaria metabólica, especialmente la que tiene que ver con la respiración.
La “especificidad” por las moléculas inorgánicas que emplean como sustratos para producir energía depende del ambiente donde se desarrolla cada especie en particular.
Otras arqueas, así como las plantas, las algas, los briófitos y las cianobacterias, son capaces de realizar fotosíntesis, es decir, que utilizan y convierten la energía lumínica de los rayos solares en energía química aprovechable.
Se ha demostrado que algunas arqueas habitan en el estómago (rumen) de algunos animales rumiantes (entre los que están las vacas, las ovejas, las cabras, etc.), por lo que estas se califican como “arqueas mutualistas”, ya que consumen parte de la fibra que ingieren estos animales y colaboran con la digestión de algunos de sus componentes.
Reproducción
Así como las bacterias, las arqueas son organismos unicelulares cuya reproducción es exclusivamente asexual. Los principales mecanismos descritos a partir de las especies mantenidas in vitro son:
– Fisión binaria, donde cada arquea se “parte” a la mitad para originar dos células idénticas
– La gemación o la “fragmentación”, donde las células desprenden “fragmentos” o “porciones” de sí que son capaces de formar células nuevas, genéticamente idénticas.
Hábitat
Las arqueas se han relacionado principalmente con ambientes “extremos”, es decir, aquellos lugares naturales que imponen serias restricciones para el desarrollo normal de los seres vivos, especialmente en términos de temperatura, pH, salinidad, anaerobiosis (ausencia de oxígeno), etc.; razón por la cual su estudio resulta sumamente interesante, pues poseen adaptaciones únicas.
Sin embargo, las técnicas más recientes de análisis molecular para la identificación de especies de microorganismos no cultivados (aislados y mantenidos in vitro en un laboratorio) han permitido detectar la presencia de arqueas en ambientes tan cotidianos como el suelo, el rumen de algunos animales, aguas oceánicas y de lagos, entre otros.
Sin embargo, la mayor parte de las arqueas que han sido identificadas en la naturaleza son clasificadas de acuerdo con el hábitat que ocupan, siendo familiares en la literatura los términos “hipertermófilos”, “acidófilos” y “termoacidófilos extremos”, “halófilos extremos” y “metanógenos”.
Los ambientes ocupados por las arqueas hipertermófilas son aquellos que se caracterizan por temperaturas constantes muy elevadas (muy por encima de las temperaturas “normales” a las cuales está sometida la mayoría de los seres vivos).
Los ambientes donde habitan los acidófilos extremos, por otra parte, son aquellos donde el pH es muy bajo y estos también pueden distinguirse por altas temperaturas (termoacidófilos extremos), entretanto los ambientes de los halófilos extremos son aquellos donde la concentración de sales es muy elevada.
Las arqueas metanógenas viven en ausencia de oxígeno o anaerobiosis, en ambientes donde pueden emplear otras moléculas como aceptores de electrones en su metabolismo y son capaces de producir metano como producto de “desecho” metabólico.
Ejemplos de especies de arqueas
Existen numerosas especies conocidas de arqueas, pero aquí solo se hará mención de algunas de ellas.
Ignicoccus hospitalis y Nanoarchaeum equitans
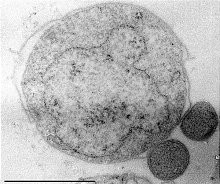
I. hospitalis pertenece al género de crenarqueotas conocido como Ignicoccus y es un organismo quimiolitoautótrofo que emplea hidrógeno molecular como donador de electrones para la reducción de azufre. Esta especie tiene el genoma más pequeño de todas las arqueas descritas in vitro hasta el momento.
I. hospitalis se comporta como un “parásito” o “simbionte” de otra especie: Nanoarchaeum equitans. Esta última no ha sido cultivada in vitro y su genoma es el más pequeño de todas las arqueas no cultivadas que han sido descritas.
Habita principalmente en ambientes marinos y no posee genes para la biosíntesis de lípidos, de aminoácidos, de nucleótidos o de cofactores, por lo que la evidencia experimental sugiere que obtiene estas moléculas gracias a su interacción con I. hospitalis.
Acidilobus saccharovorans
Esta es una especie de arquea anaeróbica termoacidófila, es decir, que vive en ambientes pobres o completamente carentes de oxígeno, con altas temperaturas y pH sumamente bajos. Fue encontrada por primera vez en cuerpos de aguas termales terrestres en Kamchatka.
Staphylothermus hellenicus
Esta arquea pertenece al filo de las crenarqueotas, específicamente al orden de los Desulfurococcales. Es una arquea heterótrofa hipertermófila (habita en ambientes muy calientes) y requiere azufre para obtener energía.