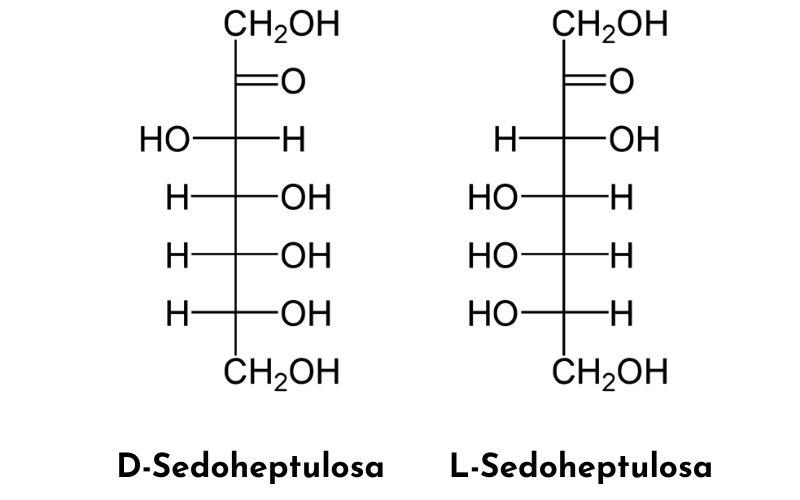
¿Qué son las heptosas?
Las heptosas son monosacáridos que poseen siete carbonos y cuya fórmula empírica es C7H14O7. Estos azúcares, como otros monosacáridos, son polihidroxilados y pueden ser: aldoheptosas, que tienen una función aldehído en el carbono uno, o cetoheptosas, que tienen un grupo cetona en el carbono 2.
Las heptosas son sintetizadas en vías metabólicas, como el ciclo de Calvin de la fotosíntesis y la fase no oxidativa de la vía de las pentosas fosfato. Son constituyentes de los lipo-polisacáridos (LPS) en la pared celular de las bacterias gramnegativas, como Escherichia coli, Klebsiella sp., Neisseria sp., Proteus sp., Pseudomonas sp., Salmonella sp., Shigella sp., y Vibrio sp.
Características de las heptosas
– Carbono. Las heptosas se caracterizan por ser monosacáridos que tener siete átomos de carbono.
– Tipos. Pueden ser aldoheptosas, con 5 carbonos asimétricos, y cetoheptosas, con 4 carbonos asimétricos.
– Fórmula química. Su fórmula química es C7H14O7.
– Estructura. Como otros monosacáridos, pueden formar estructuras cíclicas. Estas estructuras cíclicas son de gran importancia, ya que abundan en los carbohidratos naturales.
– Numerosas e importantes funciones. Las heptosas cumplen diversas funciones en procesos fundamentales de los seres vivos, como la comunicación celular y el metabolismo de carbohidratos.
Importancia biológica de las heptosas
- En la fotosíntesis y en la vía de las pentosas fosfato. En el estroma del cloroplasto se encuentran las enzimas que convierten las triosas fosfato, gliceraldehído-3-fosfato y dihidroxiacetona fosfato, producidas por la asimilación de CO₂, en almidón. La formación de triosas fosfato y la recuperación de los carbonos, para comenzar de nuevo la fijación de CO₂, constituyen dos etapas del ciclo de Calvin. Durante la etapa de recuperación de los carbonos, la enzima aldolasa se encarga de convertir a la eritrosa 4-fosfato (un metabolito de cuatro carbonos (E4P)) y a la dihidroxicetona fosfato (un metabolito de tres carbonos) en sedoheptulosa 1,7-bifosfato. Esta cetoheptosa es transformada mediante varios pasos, catalizados enzimáticamente, en ribulosa 1,5-bifosfato. La ribulosa 1,5-bifosfato es el metabolito de inicio del ciclo de Calvin. Por otra parte, la biosíntesis de sedoheptulosa 7-fosfato (S7P) tiene lugar en la vía de las pentosas fosfato, que es una vía está presente en todos los organismos vivos. En este caso, la acción de una transcetolasa transforma dos pentosas fosfato en S7P y gliceraldehído-3-fosfato (GAP). Luego, mediante dos pasos catalizados por una transaldolasa y una transcetolasa, la S7P y el GAP son transformados en fructosa-6-fosfato y GAP. Ambos son metabolitos de la glicólisis.
- En los lipo-polisacáridos (LPS) de las bacterias. Las heptosas están presentes en los lipo-polisacáridos y polisacáridos de la cápsula de las bacterias. El motivo estructural de los LPS de las enterobacterias consta de lípido A, el cual consiste en un dímero de 2-amino-2-deoxi-D-glucosa unido por enlace β-(1®6). Tiene dos ésteres de fosfato y grupos de ácidos grasos de cadena larga. El lípido A está unido a una región central mediante un puente de tres azúcares Kdo y ácido cetodeoxioctulosonico, unidos por enlaces glicosídicos (2®7). Esta región se encuentra unida a heptosas L-glicero-D-manoheptosa, con configuración anomérica alfa. Hay una región O-antigénica. Este motivo estructural está presente en bacterias gramnegativas, como Escherichia coli, Klebsiella sp., Yersinia sp., Pseudomonas sp., Salmonella sp., así como otras bacterias patógenas. Hay variantes de heptosas que incluyen diferentes configuraciones del estereocentro de las piranosas en los oligosacáridos, así como de cadenas laterales en los polisacáridos. El D-glicero-D-mano-heptopiranosil está presente en Yersinia enterocolitica, Coxiella burnetii, Mannheimia haemolitica, Aeromonas hydrophila y Vibrio salmonicida. Las heptosas D-glicero-D-mano-heptosas están presentes como unidades de cadenas laterales en la región externa de los LPS de cepas de Proteus y Haemophilus influenzae, y como cadenas laterales oligoméricas cortas unidas mediante α-(1®3) o α-(1®2), unidas al motivo estructural LPS de Klebsiella pneumoniae. En cepas de Vibrio cholerae, la región O-antigénica posee D-glicero-D-mano-heptosas con ambas configuraciones anoméricas (alfa y beta).
- En las glicoproteínas de las bacterias. Las capas de su superficie (capas S) están compuestas por subunidades de proteínas idénticas, que la cubren en una organización bidimensional. Se encuentran en las bacterias grampositivas y gramnegativas y las arqueobacterias. Las proteínas de esta capa tienen glicopéptidos, que son elongados por cadenas de polisacáridos. Las glicoproteínas de Aneurinibacillus thermoaerophilus, una bacteria grampositiva, posee unidades repetidas de disacáridos ®3)-Dglicero-β-D-mano-Hepp-(1®4)-α-L-Rhap-(1® en la capa S. Una de las funciones de las glicoproteínas es la adhesión. Por ejemplo, existe una glicoproteína que medía la adhesión como una proteína autotransportadora (AIDA-I) en cepas de E. coli. La biosíntesis de glicoproteínas ocurre mediante glicosil transferasas, como la heptosil-transferasa, que necesita ADP glicero-mano-heptosa.
Síntesis de las heptosas
La síntesis química y la combinación de métodos químicos y enzimáticos de heptosas fosfato y heptosas-nucleótido activadas han permitido elucidar las vías metabólicas que usan los microorganismos para producir estas sustancias.
Muchos métodos de síntesis preparan mano-heptosas 6-epiméricas para sintetizar L-glicero-D-mano-heptosa. Estos métodos se basan en la elongación de la cadena desde el carbono anomérico, o grupo aldehído, usando reactivos de Grignard. Las glicosilaciones son llevadas a cabo en presencia de grupos acil protectores.
De este modo, hay estereocontrol preservándose la configuración α-anomérica. Los tioglicósidos anoméricos y derivados tricloroacetimidato sirven como donadores de grupo heptosil. Los procedimientos más recientes implican la formación selectiva de β-heptósidos y derivados 6-deoxi-heptósidos.
La biosíntesis de heptosas-nucleótido activadas comienza a partir de sedoheptulosa 7-fosfato, la cual es convertida en D-glicero-D-mano-heptosa 7-fosfato. Se ha propuesto que una fosfomutasa forma el heptosil fosfato anomérico. Luego, una heptosil transferasa cataliza la formación de ADP D-glicero-D-mano-heptosa.
Por último, una epimerasa cambia la configuración del ADP D-glicero-D-mano-heptosa a ADP L-glicero-D-mano-heptosa.
Adicionalmente, se han realizado estudios químicos para conocer los mecanismos mediante los cuales estas enzimas llevan a cabo la catálisis. Por ejemplo, usan manopiranósido bencil bencilado, el cual es oxidado para dar el derivado manourónico.
El tratamiento con ácido clorhídrico transforma el derivado manourónico en diazocetona. El tratamiento con diazobencil fosfórico produce una mezcla de L-glicero-7-fosfato y D-glicero-7-fosfato.
Referencias
- Collins, P.M. Dictionary of carbohydrates with CD-ROM. Chapman & Hall/CRC.
- Nelson, D.L., Cox, M.M. Lehninger principles of biochemistry. W. H. Freeman.
- Pigman, W. The carbohydrates: chemistry, biochemistry, physiology. Academic Press.
- Stick, R.V., Williams, S.J. Carbohydrates: the essential molecules of life. Elsevier.
- Voet, D., Voet, J.G., Pratt, C.W. Fundamentals of biochemistry – life at the molecular level. Wiley.