¿Qué son las histonas?
Las histonas son proteínas básicas que interactúan con el ADN para la formación de los nucleosomas, que conforman las hebras de cromatina constituyentes de los cromosomas en los organismos eucariotas.
Los nucleosomas, complejos formados por ADN y proteínas, fueron descubiertos en 1974 y son las histonas las que ensamblan este nivel basal de organización de la cromatina. No obstante, se sabe de la existencia de las proteínas histonas desde antes de la década de los 60.
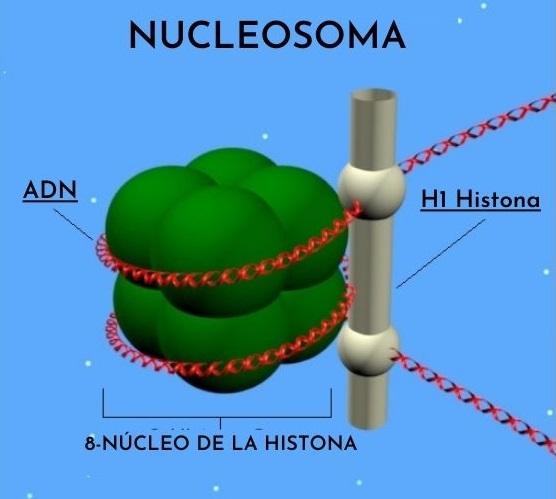
Las histonas están organizadas de tal forma que el ADN doble banda se enrolla alrededor de un centro proteico compuesto por estas proteínas que interactúan cercanamente unas con otras. El centro de histonas tiene forma de disco y el ADN da más o menos 1.7 vueltas a su alrededor.
Múltiples puentes de hidrógeno permiten la unión del ADN al centro proteico formado por las histonas en cada nucleosoma. Estos enlaces se forman, en su mayoría, entre los esqueletos aminoacídicos de las histonas y el esqueleto azúcar-fosfato del ADN. También participan algunas interacciones hidrofóbicas y enlaces iónicos.
Proteínas conocidas como “complejos remodeladores de cromatina” son las responsables de la ruptura y la formación de los enlaces de unión entre ADN e histonas, permitiendo el ingreso de la maquinaria transcripcional hacia el ADN contenido en los nucleosomas.
A pesar de la cercanía de los ácidos nucleicos con el centro proteico formado por las histonas, estas están dispuestas de manera tal que, en caso de ser necesario, permiten la entrada de factores de transcripción y otras proteínas relacionadas con la expresión o el silenciamiento genético.
Las histonas pueden sufrir diversas modificaciones que generan múltiples variantes, haciendo posible la existencia de muchas formas de cromatina diferentes que tienen la propiedad de modular de distintas formas la expresión genética.
Características de las histonas
- Son de las proteínas eucariotas más conservadas en la naturaleza. Se ha demostrado, por ejemplo, que la histona H4 del guisante difiere en tan solo dos de las 102 posiciones aminoacídicas de la proteína H4 de vaca.
- Las histonas son proteínas relativamente pequeñas, con no más de 140 aminoácidos. Son ricas en residuos aminoacídicos básicos, por lo que tienen una carga neta positiva, lo que contribuye a su interacción con el ácido nucleico, de carga negativa, para formar los nucleosomas.
- Hay histonas nucleosomales y de unión o puente. Las histonas nucleosomales son H3, H4, H2A y H2B, mientras que las histonas de unión pertenecen a la familia de las histonas H1.
- Durante el ensamblaje del nucleosoma se forman inicialmente los dímeros específicos H3-H4 y H2A-H2B. Dos dímeros H3-H4 se unen después para formar tetrámeros que posteriormente se combinan con los dímeros H2A-H2B, formando el centro octamérico.
- Todas las histonas son sintetizadas principalmente durante la fase S del ciclo celular, y los nucleosomas son ensamblados en las hélices de ADN nacientes, justo después de la horquilla de replicación.
Estructura
La estructura general de las histonas incluye una región aminoacídica básica y una región carboxilo globular sumamente conservada entre los organismos eucariotas.
Un motivo estructural conocido como “pliegue de histonas”, compuesto por tres hélices alfa conectadas por dos horquillas y que conforman un pequeño centro hidrofóbico, es el responsable de las interacciones proteína-proteína entre las histonas que forman el nucleosoma.
Es este pliegue de histonas el que conforma el dominio carboxilo globular de dichas proteínas nucleosomales en todos los eucariotas.
Las histonas también tienen pequeñas “colas” o regiones amino-terminales y otras carboxilo-terminales (accesibles a proteasas), de no más de 40 aminoácidos de longitud. Ambas regiones son ricas en aminoácidos básicos que pueden sufrir múltiples modificaciones covalentes postraduccionales.
Histonas de unión
En los eucariotas existen dos familias de histonas de unión, diferenciadas entre sí por su estructura. Unas tienen una estructura tripartita, con el dominio globular descrito anteriormente, flanqueado por unos dominios N- y C-terminales “no estructurados”, mientras que otras solo tienen un dominio C-terminal.
A pesar de que la mayor parte de las histonas se conservan, durante la embriogénesis o la maduración de las células especializadas en algunos organismos pueden surgir algunas variantes específicas. Algunas variaciones estructurales tienen que ver con modificaciones postraduccionales como las que siguen:
- Fosforilación: se piensa que está relacionada con la modificación del grado de condensación de la cromatina y se da comúnmente en residuos de serina.
- Acetilación: asociada con las regiones cromosómicas que son transcripcionalmente activas. Ocurre normalmente en las cadenas laterales de los residuos de lisina. Al ocurrir sobre estos residuos disminuye la carga positiva de los mismos, disminuyendo con ello la afinidad de las proteínas por el ADN.
- Metilación: puede darse como mono-, di- o tri- metilación de los residuos de lisina que sobresalen del núcleo proteico.
Enzimas específicas se encargan de realizar estas modificaciones covalentes en las histonas. Entre estas enzimas se incluyen las histona-acetil transferasas (HATs), los complejos histona-deacetilasas (HDACs), y las histona-metiltransferasas y demetilasas.
Tipos
La caracterización de las histonas ha sido realizada por diversas técnicas bioquímicas, entre las que destacan las cromatografías que se basan en resinas de intercambio catiónico débil.
Ciertos autores establecen una forma de clasificación en el que se distinguen 5 tipos principales de histonas en los eucariotas: FI, con proteínas de 21 kDa; F2A1 o FIV, de más o menos 11.3 kDa; F2A2 o FIIbI, 14.5 kDa; F2B o FIIb2, con un peso molecular de 13.7 kDa, y F3 o FIII, de 15.3 kDa.
Todos estos tipos de histonas, con la excepción del grupo de las FI, se encuentran en cantidades equimolares en la células.
Otra clasificación, con la misma validez y quizá la más empleada en la actualidad, propone la existencia de dos tipos de histonas diferentes, a saber: las que forman parte del octámero del nucleosoma y las histonas de unión o puente, que unen los nucleosomas entre sí.
Algunas variantes pueden darse también entre especies, y a diferencia de las histonas de núcleo, las variantes son sintetizadas durante la interfase y son insertadas en la cromatina preformada a través de un proceso dependiente de la energía liberada de la hidrólisis del ATP.
Histonas nucleosomales
El centro de un nucleosoma consiste en un par de cada una de las cuatro histonas constituyentes: H2a, H2b, H3 y H4; sobre las cuales se enrollan segmentos de ADN de alrededor de 145 pares de bases.
Las histonas H4 y H2B son, en principio, invariables. Algunas variaciones son evidentes, sin embargo, en las histonas H3 y H2A, cuyas propiedades biofísicas y bioquímicas alteran la naturaleza normal del nucleosoma.
Una variante de la histona H2A en humanos, la proteína H2A.Z posee una gran región acídica y puede favorecer la estabilidad del nucleosoma dependiendo de las variantes de la histona H3 con las que se asocie.
Estas histonas muestran cierta variabilidad entre especies, siendo un caso especial el de la histona H2B, para la cual el primer tercio de la molécula es altamente variable.
Histonas de unión
Las histonas de unión o de puente son las histonas de la clase H1. Estas son las responsables de la unión entre nucleosomas y de la protección del ADN que sobresale al comienzo y al final de cada partícula.
A diferencia de las histonas nucleosomales, no todas las histonas de tipo H1 poseen la región globular del “pliegue” de histonas. Estas proteínas se unen al ADN entre nucleosomas, facilitando un cambio en el equilibrio de la cromatina hacia un estado más condensado y menos activo, transcripcionalmente hablando.
Estudios han relacionado estas histonas con el envejecimiento, reparación del ADN y procesos apoptóticos, por lo que se piensa que las mismas tienen un papel crucial en el mantenimiento de la integridad genómica.
Funciones
Todos los residuos aminoacídicos de las histonas participan, de una u otra manera, en su interacción con el ADN, lo que explica el hecho de que las mismas estén tan conservadas entre los reinos de organismos eucariotas.
La participación de las histonas en el empaquetamiento del ADN en forma de cromatina es de gran relevancia para los organismos multicelulares complejos, en los que los diferentes linajes celulares pueden especializarse tan solo cambiando la accesibilidad de sus genes a la maquinaria transcripcional.
Las regiones genómicas transcripcionalmente activas son densas en nucleosomas, lo que sugiere que la asociación del ADN con las proteínas histonas es crucial para la regulación negativa o positiva de su transcripción.
Asimismo, a lo largo de la vida de una célula, la respuesta a gran número de estímulos, tanto internos como externos, depende de pequeños cambios en la cromatina, que normalmente tienen que ver con remodelación y modificación postraduccional de las histonas que se encuentran en estrecha relación con el ADN.
Múltiples variables de las histonas ejercen diferentes funciones en los eucariotas. Una de ellas tiene que ver con la participación de una variante de la histona H3 en la formación de las estructuras centroméricas responsables de la segregación de los cromosomas durante la mitosis.
Se ha demostrado que la contraparte de esta proteína en otros eucariotas es esencial para el ensamblaje del cinetocoro proteico, al cual se unen los microtúbulos del huso durante la mitosis y la meiosis.
Referencias
- Harvey, A. C., & Downs, J. A. What functions do linker histones provide? Molecular Microbiology.
- Henikoff, S., & Ahmad, K. Assembly of Variant Histones into Chromatin. Annu. Rev. Cell. Dev. Biol.
- Isenberg, I. Histones. Annu. Rev. Biochem.