¿Qué es la teoría endosimbiótica?
La teoría endosimbiótica, o endosimbiosis (endo, dentro, y simbiosis, vivir juntos), plantea el origen de las distintas organelas presentes en los eucariotas a partir de relaciones simbióticas entre procariotas ancestrales.
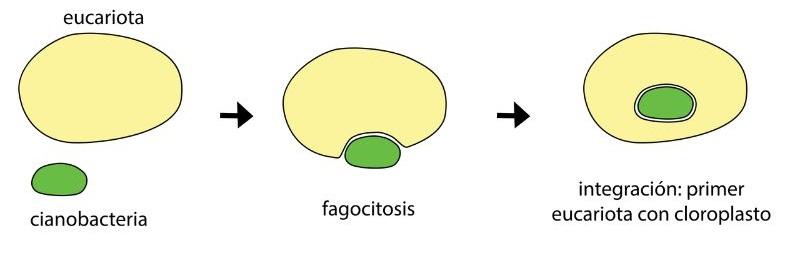
En esta relación, un procariota fue engullido por uno de mayor tamaño. Eventualmente, el organismo más pequeño no fue digerido, sino que sobrevivió y se incorporó al citoplasma de su huésped. Para que ocurriese la continuidad evolutiva, ambos organismos debieron tener tiempos de replicación similares. La consecuencia de esta división sincronizada es que la descendencia del huésped contenía el organismo simbionte.
Así, se plantea que los cloroplastos son el resultado de una endosimbiosis entre un organismo heterótrofo y una cianobacteria que, con el tiempo, pasó a ser un plastidio. Del mismo modo, se especula que las mitocondrias tienen su origen evolutivo en el grupo de las alfa-proteobacterias.
Aunque estas ideas empezaron a formarse en el siglo XIX, fueron retomadas, modificadas y sustentadas adecuadamente a mediados de los años 60 por Lynn Margulis.
Algunas de las evidencias de esta teoría son la similitud entre las organelas y las bacterias en términos de tamaño, organización del genoma, estructura de los ribosomas y homología moleculares.
Historia de la teoría endosimbiótica
1900-1920: contribuciones de Mereschkowsky, Portier y Wallien
Las primeras ideas relacionadas con teorías endosimbióticas se le atribuyen al botánico ruso Constantin Mereschkowsky, quien describió el origen de los plastidios (organelas de las plantas que incluyen cloroplastos, cromoplastos, amiloplastos, entre otros) en 1905.
El planteamiento de este autor consistió básicamente en un evento simbiótico entre una cianobacteria “reducida” y un huésped. Si bien el pensamiento del científico Mereschkowsky resultó innovador para la época, no daba una explicación para el origen de las demás organelas eucariotas.
En 1918, el biólogo francés Paul Portier notó cierto parecido entre las bacterias y las mitocondrias. Aunque las ideas iban por buen camino, el autor propuso el cultivo de mitocondrias fuera de las células, metodología rechazada por sus colegas.
A mediados de los años 20, el origen de las mitocondrias fue extendido por el biólogo estadounidense Ivan Wallin, convencido de que estas organelas eran descendientes de organismos procariotas. Pero no encontró un mecanismo plausible para la transformación planteada, por lo que la teoría endosimbiótica fue olvidada.
1960: contribuciones de Lynn Margulis
En los años 60, la joven investigadora de la Universidad de Boston, Lynn Margulis, propuso la teoría endosimbiótica de manera bastante robusta, basada en evidencia citológica, bioquímica y paleontológica.
Actualmente, la teoría endosimbiótica es aceptada con normalidad, pero en los tiempos de Margulis sus ideas fueron tratadas con marcado escepticismo, lo que causó el rechazo de su trabajo por más de 15 revistas científicas.
¿Qué propone la teoría endosimbiótica?
El origen de la primera célula procariota data de hace más de 3.5 mil millones de años, y permaneció de esta manera durante aproximadamente 1.5 mil millones de años adicionales. Pasado este lapso, se presume que aparecieron las primeras células eucariotas, identificadas por su complejidad, presencia de núcleo y de organelas.
En biología, uno de los tópicos más importantes es el origen y evolución de la célula eucariota, y una de las teorías que busca explicarlo es la teoría endosimbiótica.
Esta propone el origen de los organelos a partir de eventos de simbiosis entre organismos procariotas ancestrales, que con el paso del tiempo se integraron, el organismo engullido se redujo y pasó a ser un componente citoplasmático del más grande.
Una de las consecuencias de la teoría endosimbiótica es la transferencia horizontal de genes entre los organismos procariotas involucrados en la asociación simbiótica, desde la nueva “organela” hasta el genoma nuclear del huésped.
Evidencias que sustentan la teoría endosimbiótica
- Tamaño. El tamaño de las organelas eucariotas (llámense cloroplastos o mitocondrias) es bastante similar al de los organismos bacterianos actuales.
- Ribosomas. Una de las diferencias más notorias entre los linajes eucariotas y procariotas reside en el tamaño de las subunidades grandes y pequeñas que componen los ribosomas (estructuras implicadas en la síntesis de proteínas). Los cloroplastos y las mitocondrias poseen en su interior ribosomas y estos exhiben los rasgos de los ribosomas descritos en las eubacterias.
- Material genético. Tanto los cloroplastos como las mitocondrias se caracterizan por presentar su propio genoma circular, tal como los organismos procariotas.
- Genoma de la mitocondria. El genoma de la mitocondria está formado de los genes que codifican para un pequeño número de ARN ribosomal y ARN de transferencia, implicados en la síntesis de proteínas del organelo en cuestión. La inmensa mayoría de proteínas estructurales y enzimas mitocondriales son codificadas por genes que residen en el material genético nuclear. La organización del genoma mitocondrial varía extensamente entre los linajes de eucariotas. En el humano, por ejemplo, el genoma mitocondrial es una molécula circular que alberga unos 16.569 pares de bases que codifican para dos ARN ribosomales, 22 ARN de transferencia y solamente 13 proteínas.
- Genoma de los cloroplastos. En contraste con el genoma mitocondrial, el de los cloroplastos es un poco más grande y contiene la información necesaria para la síntesis de aproximadamente 120 proteínas.
- Consecuencias del genoma de los orgánulos. El modo de división peculiar de los cloroplastos y mitocondrias muestra un patrón no mendeliano. Es decir, la reproducción ocurre por medio de la reproducción del mismo (como en las bacterias) y no por síntesis celular de novo. Este fenómeno ocurre por la existencia de material genético único que no encontramos en el núcleo de la célula. Las mitocondrias son heredadas por línea materna y en la gran mayoría de las plantas con reproducción sexual los cloroplastos son aportados al cigoto en formación por la planta madre.
- Homologías moleculares. Gracias al secuenciamiento de genes, ha quedado en evidencia que las secuencias del ARN ribosómico y de los demás genes en mitocondrias y cloroplastos están más relacionadas con las secuencias de las bacterias que con las albergadas en el núcleo de los eucariotas. Las secuencias del ADN de las mitocondrias son marcadamente parecidas a las secuencias encontradas en un grupo puntual de bacterias denominadas alfa-proteobacterias. Esta evidencia apunta a que el posible organismo que participó en el evento endosimbiótico era una alfa-proteobacteria ancestral. En contraste, las secuencias de los cloroplastos parecen estar estrechamente relacionadas con las cianobacterias, un grupo de eubacterias con la maquinaria enzimática necesaria para llevar a cabo las reacciones fotosintéticas.
Origen de la mitocondria
La mitocondria actual pudo haber surgido por un evento ocurrido entre 1 y 1.5 mil millones de años, donde una célula anaeróbica de gran tamaño engulló una bacteria aeróbica más pequeña, con la maquinaria enzimática necesaria para la fosforilación oxidativa.
El organismo aeróbico le proporcionaba a su huésped la capacidad de generar más ATP por cada molécula orgánica degradada.
A medida que la teoría endosimbiótica ganó aceptación en la comunidad científica, la identidad taxonómica de los organismos ancestrales involucrados en la simbiosis fue arduamente discutida.
Hoy en día, se maneja la idea de que el huésped de gran tamaño era una arquea y el organismos engullido, como se dijo, una alfa-proteobacteria, aunque algunas variantes de la teoría proponen una bacteria anaeróbica, ya que existen varias formas anaeróbicas de mitocondrias, como los hidrogenosomas.
Origen de los plastidios
- Plastidios primarios. Los rápidos avances en las técnicas de secuenciación y procesamiento bioinformático, en los 90, arrojaron evidencia a nivel molecular. Los estudios comparativos basados en filogenias moleculares lograron trazar el origen de los genes codificados por los plastidios de las plantas hasta las cianobacterias. Además, demostraron la transferencia de genes desde el genoma del endosimbionte hasta el genoma nuclear del huésped. Se calcula que el establecimiento de los primeros plastidios ocurrió hace 1.5 mil millones de años, aunque la cifra temporal sigue siendo controversial entre los científicos.
- Plastidios secundarios. Si bien el establecimiento de una cianobacteria ancestral dentro de un huésped procariota explica el origen de los plastidios primarios, la historia evolutiva se torna compleja cuando se busca explicar el origen de los plastidios secundarios encontrados en algunos organismos fotosintéticos. Estos plastidios se caracterizan por la presencia de membranas adicionales, es decir, una o dos membranas extras a las dos que usualmente rodean la organela. Este número de membranas complica la interpretación, pues si un procariota ancestral engulló una cianobacteria, no obtendría las membranas adicionales. Por ello, una respuesta a este problema evolutivo fue proponer múltiples eventos endosimbióticos. En este contexto, una célula de planta con un plastidio ya existente fue engullida por un segundo huésped, y eventualmente se redujo a un plastidio. En algunos casos se plantea un tercer evento endosimbiótico. El número de simbiosis y la naturaleza del huésped son discutidos en la literatura, aunque existe evidencia empírica que sustenta la idea de múltiples eventos endosimbióticos.
Referencias
- Anderson, P. W. Suggested model for prebiotic evolution: The use of chaos. Proceedings of the National Academy of Sciences.
- Chan, C. X. & Bhattacharya, D. The Origin of Plastids. Nature Education.
- Gama, M. Biología 1: un Enfoque Constructivista. Pearson Educación.
- Gray, M. W. Lynn Margulis and the endosymbiont hypothesis: 50 years later. Molecular biology of the cell.
- Margulis, L. Serial endosymbiotic theory (SET) and composite individuality. Microbiology Today.