¿Qué es el ciclo del glioxilato?
El ciclo del glioxilato es una ruta metabólica presente en las plantas, en algunos microorganismos y en los animales invertebrados (ausente en todos los vertebrados), a través de la cual dichos organismos pueden convertir las grasas en hidratos de carbono (azúcares).
Esta ruta fue descubierta en 1957, mientras Kornberg, Krebs y Beevers trataban de dilucidar cómo las bacterias como Escherichia coli podían crecer en presencia de acetato como única fuente de carbono, y cómo las plántulas en germinación del tártago (Ricinus communis) podían convertir las grasas en carbohidratos.
Los estudios de estos tres investigadores llevaron al descubrimiento de dos enzimas, conocidas como isocitrato liasa y malato sintasa, que, junto con las enzimas del ciclo de Krebs, permiten la síntesis de succinato a partir de dos moléculas de acetil-coA.
El succinato así producido es convertido a malato a través del ciclo de ácidos tricarboxílicos, pudiendo posteriormente emplearse para la producción de glucosa por medio de la gluconeogénesis.
Esta ruta ocurre, en las plantas, en unos orgánulos especiales denominados glioxisomas y es fundamental para la supervivencia de las plántulas durante las primeras etapas de la germinación.
Características del ciclo de glioxilato
– Presente en algunos organismos. El ciclo del glioxilato se encuentra en plantas (en los glioxisomas), bacterias y hongos. En las plantas, es clave durante la germinación de las semillas que almacenan lípidos, ya que convierte las reservas de grasa en carbohidratos que alimentan el crecimiento del brote.
– Conversión de ácidos grasos en carbohidratos. A diferencia del ciclo de Krebs, este ciclo permite la síntesis neta de glucosa a partir de moléculas de acetil-CoA derivadas de ácidos grasos, algo que el ciclo de Krebs no puede hacer por sí solo. Es posible porque evita las reacciones de descarboxilación que eliminan carbono en el ciclo de Krebs, permitiendo que los carbonos se usen para generar glucosa.
– Enzimas. La isocitrato liasa convierte el isocitrato en glioxilato y succinato. El glioxilato es un intermediario clave que no está presente en el ciclo de Krebs. Por su parte, la malato sintasa combina el glioxilato con una molécula de acetil-CoA para formar malato, el cual luego puede ser utilizado para sintetizar glucosa a través de la gluconeogénesis.
– Modificación del ciclo de Krebs. Comparte varias enzimas con el ciclo de Krebs, pero evita las reacciones que liberan CO₂, lo que permite la retención de carbono necesario para la síntesis de carbohidratos.
– Ubicación. En las plantas, este ciclo tiene lugar en unos orgánulos llamados glioxisomas, especializados en la conversión de lípidos en carbohidratos.
– Importante para la germinación de semillas. En plantas, el ciclo es especialmente relevante en las semillas oleaginosas (que almacenan grasas), como el girasol o el ricino. Durante la germinación, las grasas se convierten en carbohidratos que son usados para el crecimiento de las plántulas hasta que puedan realizar la fotosíntesis.
– Precursores biosintéticos. Además de proporcionar glucosa, el ciclo del glioxilato produce intermediarios como el succinato, que puede ser utilizado en otras rutas biosintéticas importantes.
Reacciones en el ciclo del glioxilato
El acetato proveniente de la degradación de los ácidos grasos funciona como combustible rico en energía y como una fuente de fosfoenolpiruvato para la síntesis de glucosa a través de la gluconeogénesis. El proceso se da como sigue:
- Pasos del ciclo del glioxilato
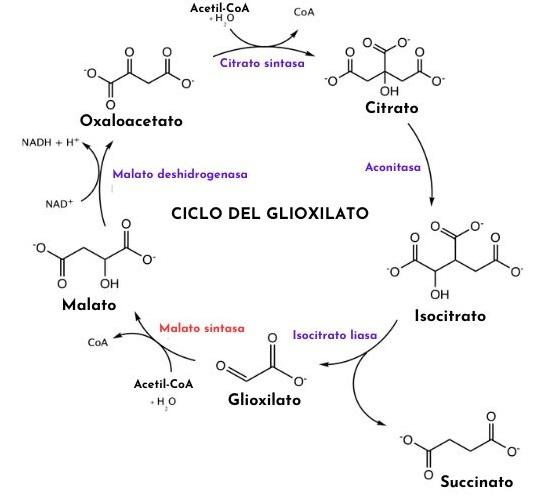
- 1. La ruta del glioxilato, de forma similar a la del ciclo de Krebs, comienza con la condensación de una molécula de acetil-CoA con otra de oxalacetato para rendir citrato, reacción catalizada por la enzima citrato sintasa.
- 2. La enzima aconitasa convierte este citrato en isocitrato.
- 3. El isocitrato es empleado como sustrato de la enzima isocitrato liasa para formar los compuestos succinato y glioxilato.
-
- 4. El glioxilato es tomado por la enzima malato sintasa para producir malato a través de su condensación con una segunda molécula de acetil-CoA.
- 5. El malato es convertido en oxalacetato por la malato deshidrogenasa y dicho compuesto puede servir de precursor para la ruta gluconeogénica o condensarse con otro acetil-CoA para reiniciar el ciclo una vez más.
- 6. El succinato producido también puede ser convertido a fumarato y este a malato, aportando mayor cantidad de moléculas de oxalacetato para la formación de glucosa. De lo contrario, esta molécula también puede ser exportada hacia la mitocondria para funcionar en el ciclo de Krebs. El oxalacetato entra a la ruta gluconeogénica para la producción de glucosa gracias a su conversión en fosfoenolpiruvato, que es catalizada por la enzima fosfoenolpiruvato carboxiquinasa.
Regulación del ciclo del glioxilato
Puesto que los ciclos del glioxilato y de los ácidos tricarboxílicos comparten numerosos intermediarios entre sí, existe una coordinada regulación entre ambos.
Además, es necesario que existan mecanismos de control, pues la síntesis de glucosa y de otras hexosas a partir del acetil-CoA (proveniente de la degradación de grasas) implica la participación de al menos cuatro rutas:
- La β-oxidación de ácidos grasos que produzca las moléculas de acetil-CoA necesarias tanto para el ciclo del glioxilato como para el ciclo de Krebs y que, en las plantas, tiene lugar en los glioxisomas.
- El ciclo del glioxilato, que también ocurre en los glioxisomas y que, como se dijo, produce intermediarios como el succinato, malato y oxalacetato.
- El ciclo de Krebs, que tiene lugar en la mitocondria y en el cual también se producen los intermediarios succinato, malato y oxalacetato.
- La gluconeogénesis, que ocurre en el citosol y que contempla el uso del oxalacetato convertido en fosfoenolpiruvato para sintetizar glucosa.
El principal punto de control está en la enzima isocitrato deshidrogenasa, cuya regulación implica una modificación covalente por adición o remoción de un grupo fosfato.
Cuando la enzima es fosforilada se inactiva, por lo que el isocitrato se dirige hacia la vía del glioxilato para la producción de glucosa.
Funciones del ciclo del glioxilato
- Conversión de ácidos grasos en carbohidratos. Su función más importante es permitir la síntesis de carbohidratos (glucosa) a partir de ácidos grasos. Esto es esencial en situaciones donde los carbohidratos son escasos, y los organismos necesitan utilizar sus reservas de grasa como fuente de energía. El ciclo del glioxilato, al evitar las reacciones de descarboxilación del ciclo de Krebs, retiene carbonos que luego pueden utilizaarse para la gluconeogénesis (síntesis de glucosa).
- Aporte de energía en la germinación de semillas. En las plantas, el ciclo del glioxilato es clave durante la germinación de las semillas oleaginosas. Durante la germinación, estas plantas convierten sus reservas de lípidos en carbohidratos para proporcionar energía a la plántula en crecimiento, hasta que comience la fotosíntesis.
- Supervivencia en condiciones de escasez. En microorganismos, como bacterias y hongos, el ciclo del glioxilato permite el crecimiento y supervivencia en ambientes donde hay poca disponibilidad de azúcares. Estos organismos pueden utilizar ácidos grasos u otros compuestos derivados de lípidos como fuente de carbono para generar glucosa y otros compuestos esenciales.
- Mantenimiento de la homeostasis del carbono. Al permitir el uso de ácidos grasos como fuente de carbono, el ciclo del glioxilato juega un papel en la homeostasis del carbono, manteniendo el equilibrio de fuentes de carbono en el organismo, especialmente cuando los carbohidratos no están disponibles.
- Promueve la gluconeogénesis. Proporciona intermediarios, como el malato y el oxalacetato, esenciales para la gluconeogénesis, el proceso mediante el cual se sintetiza glucosa a partir de precursores no carbohidratos.
Referencias
- Dey, P., Harborne, J. Plant Biochemistry. Academic Press.
- Ensign, S.A. Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation. Molecular Microbiology.
- Garrett, R., Grisham, C. Biochemistry (4th ed.). Brooks/Cole. CENGAGE Learning.
- Mathews, C., van Holde, K., Ahern, K. Biochemistry (3th ed.). Pearson.
- Rawn, J.D. Biochemistry. Neil Patterson Publishers.