¿Qué es la especiación parapátrica?
La especiación parapátrica propone el surgimiento de nuevas especies a partir del aislamiento reproductivo de dos subpoblaciones que se encuentran una al lado de la otra. Es uno de los tres modelos básicos de especiación, y se ajusta a un estado “intermedio” entre los modelos alopátricos y simpátricos.
Esta teoría implica la especiación en poblaciones distribuidas en áreas contiguas y donde existe un flujo moderado de genes entre ambas regiones. Al existir cierto grado de aislamiento entre las dos subpoblaciones, cada una de estas puede incrementar los niveles de independencia genética.
Con el pasar del tiempo, las especies pueden desarrollar mecanismos de aislamiento reproductivo y el proceso de especiación se habrá completado.
Especiación: formación de nuevas especies
Es frecuente que cualquier tema de discusión en la biología evolutiva empiece con las contribuciones del famoso naturalista británico Charles Darwin.
En su obra maestra, El origen de las especies, Darwin propone el mecanismo de la selección natural, y postula —entre otras cosas— cómo las nuevas especies pueden formarse por acción gradual de este mecanismo, durante largos periodos de tiempo.
Pero, ¿qué es una especie? Esta interrogante ha sido de gran estudio y controversia para los biólogos. Aunque existen decenas de definiciones, el concepto mayormente usado y aceptado es el concepto biológico de especie, formulado por Ernst Mayr.
Para Mayr, una especie se define como: “grupos de poblaciones naturales entrecruzables que se encuentran reproductivamente aislados de otros grupos”. Un punto crítico en esta definición es el aislamiento reproductivo entre los grupos que llamamos especies.
De este modo, una nueva especie se forma cuando individuos pertenecientes a dos poblaciones divergentes no se reconocen entre sí como parejas potenciales.
Modelos de especiación
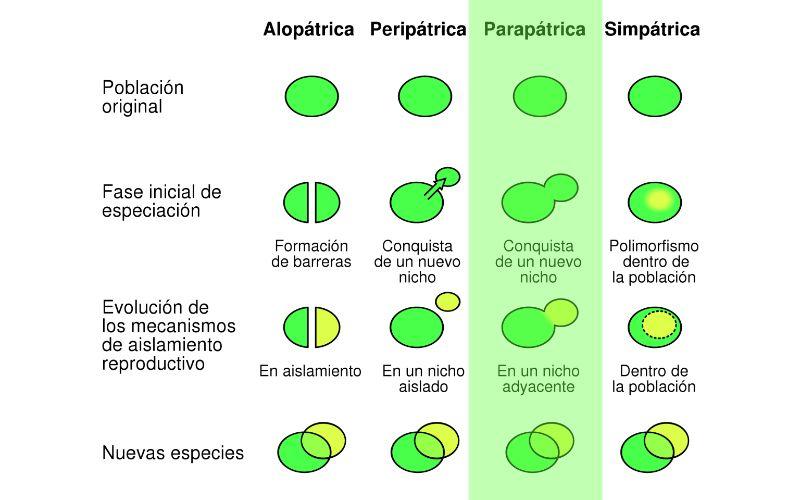
Dependiendo del contexto geográfico en donde ocurra la especiación, los autores usan un sistema de clasificación que incluye tres modelos principales: especiación alopátrica, simpátrica y parapátrica.
Si el origen de las nuevas especies involucra aislamiento geográfico total (por surgimiento de una barrera geográfica, como un río o una montaña), la especiación es alopátrica. Si las especies se forman en una misma área geográfica sin separación alguna, es especiación simpátrica.
Un modelo intermedio es la especiación parapátrica, donde las nuevas especies surgen en regiones geográficas continuas. A continuación describiremos en detalle este modelo intermedio.
Es importante mencionar que la distinción entre los tres tipos de especiación puede que no sea claro y se solapen entre sí.
Proceso de especiación parapátrico
En la especiación parapátrica ocurre la división de dos “subpoblaciones” biológicas que se ubican una al lado de la otra, sin ninguna barrera geográfica que impida el flujo de genes entre ambos demos (un “demo” es otro término ampliamente usado en la literatura para referirse a las poblaciones).
La especiación parapátrica puede ocurrir de la siguiente manera: inicialmente, una población se encuentra distribuida homogéneamente en cierta zona geográfica. Con el paso del tiempo, la especie evoluciona un patrón de “clina”.
Este modelo clinal fue propuesto por Fisher en 1930. Aunque es el modelo tradicional, existen otras propuestas —como la especiación stepping-stone—.
Modelo clinal
Una clina es un gradiente fenotípico que ocurre en una misma especie (por ejemplo, en términos de tamaño corporal: los individuos se distribuyen desde tallas grandes hasta tallas pequeñas).
El origen de la clina puede ocurrir por un cambio geográfico abrupto. Gracias al cambio, algunas formas logran adaptarse a las condiciones de un lado, mientras que la población restante se adapta al otro lado.
Entre cada uno de los límites, se formaría una zona híbrida, donde los integrantes de cada lado del nuevo gradiente geográfico entran en contacto y hay flujo génico entre ambas subpoblaciones. Sin embargo, ahora las especies de cada “lado” pueden ser reconocidas como entes separados.
Estas dos formas pueden recibir nombres taxonómicos diferentes, pudiendo ser clasificadas como razas o bien como subespecies.
Zona de tensión
Puede que en la zona híbrida se forme una zona de tensión, que favorece el proceso de especiación. En esta zona, la formación de los híbridos es desventajosa, es decir, los híbridos tienen una adecuación biológica menor que las especies parentales.
Supongamos que un individuo es homocigoto dominante para cierto rasgo (AA), y está adaptado hacia un lado de la zona geográfica. Al otro lado, se encuentran los individuos homocigotos recesivos (aa), adaptados a dicha región.
Si ocurre un cruce en la zona híbrida entre las dos “razas” o “subespecies” y el híbrido entre ambas (en este caso, el heterocigoto Aa) tiene una menor adecuación biológica o fitness, es una zona de tensión. Según las evidencias empíricas, casi todas las zonas híbridas conocidas entran en la definición de zona de tensión.
Así, la selección natural favorecerá el apareamiento selectivo entre cada una de las variantes que viven en las regiones geográficas continuas. Es decir, los de la izquierda se reproducirán entre sí y lo mismo ocurrirá en el lado derecho.
Evidencia
Aunque las bases teóricas de la especiación parapátrica la convierte en un modelo posible y atractivo, la evidencia es relativamente poca y no es contundente.
No existen suficientes pruebas que logren ilustrar cada paso del proceso. Sin embargo, el modelo no está descartado del todo y puede que en ciertos casos suceda.
Ejemplos de especiación parapátrica
Especiación en la gramínea de la especie Anthoxanthum odoratum
La gramínea Anthoxanthum odoratum perteneciente a la familia Poaceae, representa un ejemplo muy ilustrativo de especiación parapátrica.
Algunas de estas plantas viven en zonas donde el suelo se encuentra contaminado por una variedad de metales pesados. De esta forma, solamente las variantes de la gramínea que son capaces de tolerar la contaminación pueden creer en dichas regiones.
En contraste, las plantas vecinas que no viven en suelos contaminados no han experimentado un proceso de selección hacia la tolerancia a los metales pesados.
Las formas tolerante y no tolerante están lo suficientemente cerca como para fertilizarse entre sí (requisito para que el proceso de especiación pueda ser considerado como parapátrico). Sin embargo, ambos grupos han desarrollado tiempos de floración diferentes, estableciendo una barrera temporal para el flujo de genes.
Cuervos de la especie Corvus corone y C. cornix
Estas dos especies de cuervos se distribuyen por Europa y son un ejemplo clásico de una zona híbrida. C. corvix se ubica más hacia el este, mientras que su compañero se localiza hacia el oeste, con un punto de encuentro de ambas especies en Europa central.
Aunque cada especie tiene características fenotípicas propias, en la zona donde se cruzan pueden producir híbridos. El cruce es una señal de que el proceso de especiación entre los dos cuervos aún no se ha completado y el aislamiento reproductivo no está completamente establecido.
Insectos de la fruta
Los insectos suelen poner sus huevos en el mismo tipo de frutas donde nacieron, de modo que se especializan en aparearse, alimentarse y reproducirse en ese mismo tipo de fruta. Eso hace que haya menor flujo genético, y los insectos de cada tipo de fruta queden aislados, formándose nuevas especies.
Los pinzones de Darwin
Los pinzones que observó Darwin en las islas Galápagos tenían picos pequeños y grandes. Se dio cuenta de que obedecía a la dieta que consumían: los de pico pequeño comían semillas pequeñas, y los de picos grandes, semillas grandes. A pesar de estar en la misma zona geográfica, la dieta determinó el surgimiento de dos especies.
Referencias
- Dieckmann, U., Doebeli, M., Metz, J. A., & Tautz, D. Adaptive speciation. Cambridge University Press.
- Gavrilets, S. Fitness landscapes and the origin of species. Princeton University Press.